Archives
- 2026-07
- 2026-06
- 2026-05
- 2026-04
- 2026-03
- 2026-02
- 2026-01
- 2025-12
- 2025-11
- 2025-10
- 2025-03
- 2025-02
- 2025-01
- 2024-12
- 2024-11
- 2024-10
- 2024-09
- 2024-08
- 2024-07
- 2024-06
- 2024-05
- 2024-04
- 2024-03
- 2024-02
- 2024-01
- 2023-12
- 2023-11
- 2023-10
- 2023-09
- 2023-08
- 2023-07
- 2023-06
- 2023-05
- 2023-04
- 2023-03
- 2023-02
- 2023-01
- 2022-12
- 2022-11
- 2022-10
- 2022-09
- 2022-08
- 2022-07
- 2022-06
- 2022-05
- 2022-04
- 2022-03
- 2022-02
- 2022-01
- 2021-12
- 2021-11
- 2021-10
- 2021-09
- 2021-08
- 2021-07
- 2021-06
- 2021-05
- 2021-04
- 2021-03
- 2021-02
- 2021-01
- 2020-12
- 2020-11
- 2020-10
- 2020-09
- 2020-08
- 2020-07
- 2020-06
- 2020-05
- 2020-04
- 2020-03
- 2020-02
- 2020-01
- 2019-12
- 2019-11
- 2019-10
- 2019-09
- 2019-08
- 2019-07
- 2019-06
- 2019-05
- 2019-04
- 2018-07
-
Notably HSP expression is under
2019-10-21
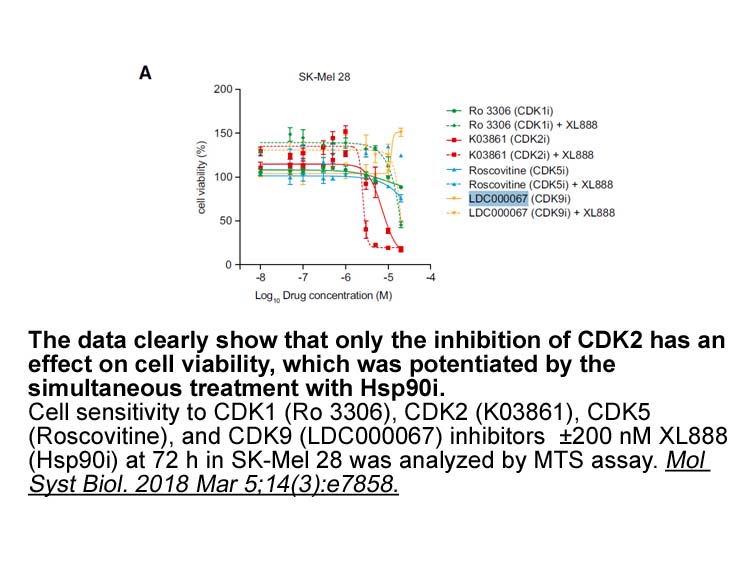
Notably, HSP27 expression is under control of the tetrameric form of CK2 holoenzyme, as knockout or knockdown of the β regulatory subunit alone reproduces the same effects of targeting the α/α′ catalytic subunits. This observation discloses the possibility to use more specific drugs that affects onl
-
It is of interest to point out that curcumin
2019-10-21
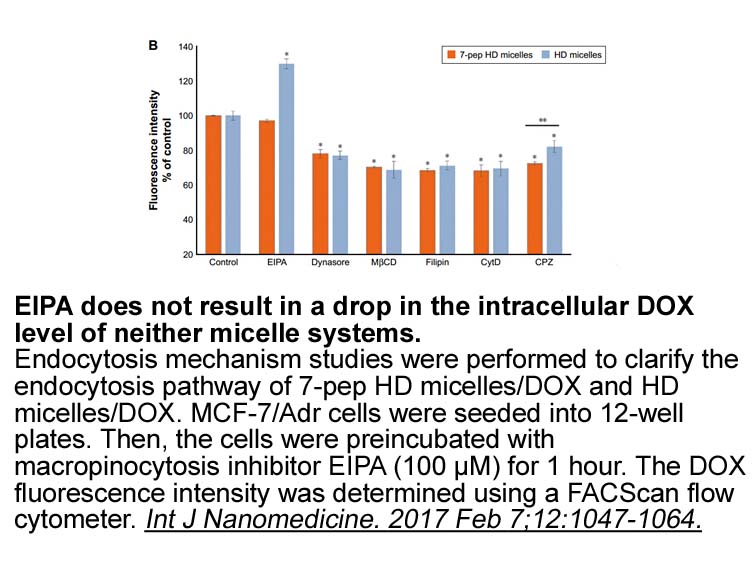
It is of interest to point out that curcumin in this study reduced body weight gain; the underlying mechanism may be due to a decrease in general Temafloxacin hydrochloride of fat, which was associated with a significantly reduced NPC1L1 expression in small intestine. Labonte et al revealed that NP
-
The mechanisms by which coregulators control the
2019-10-21
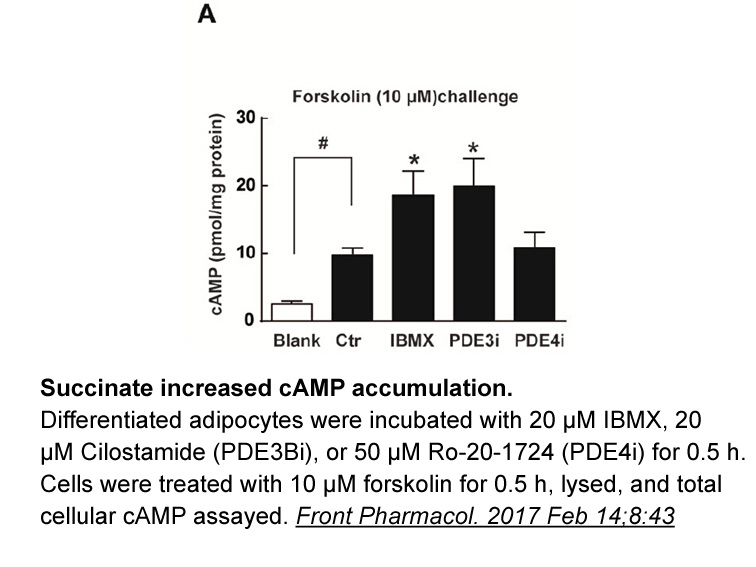
The mechanisms by which coregulators control the actions of estrogen receptors are still a topic of ongoing research. From studies in cancer cells, we have learned that a large group of coregulators have specific structural motifs that than affect their contact with ER ligand-binding domains (Heery,
-
br Materials and methods br Results br Discussion
2019-10-21
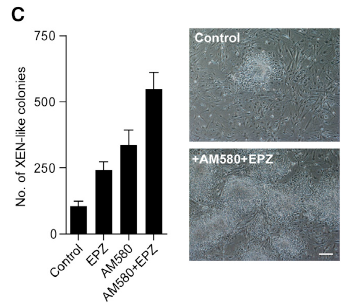
Materials and methods Results Discussion COX-2-mediated production of PGE2 is involved in cell growth and metastasis of many cancers. Previous studies indicated that COX-2 was overexpressed in many cancer tissues and that PGE2 increased cancer cell growth, a process that could be suppressed
-
The prospects of the use of this novel approach for
2019-10-21
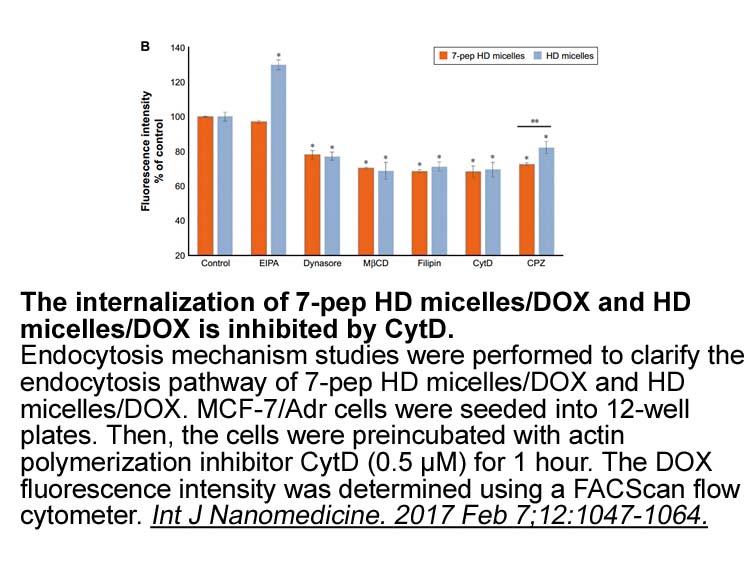
The prospects of the use of this novel approach for the selective local thermo-activation of enzymes include biomedical and biotechnological applications. As NPs could be engineered to gain access to LFM-A13 australia through the endosomal compartment [47] or through non-endocytotic pathways [48], [
-
br Transparency document br Introduction Cigarette
2019-10-21
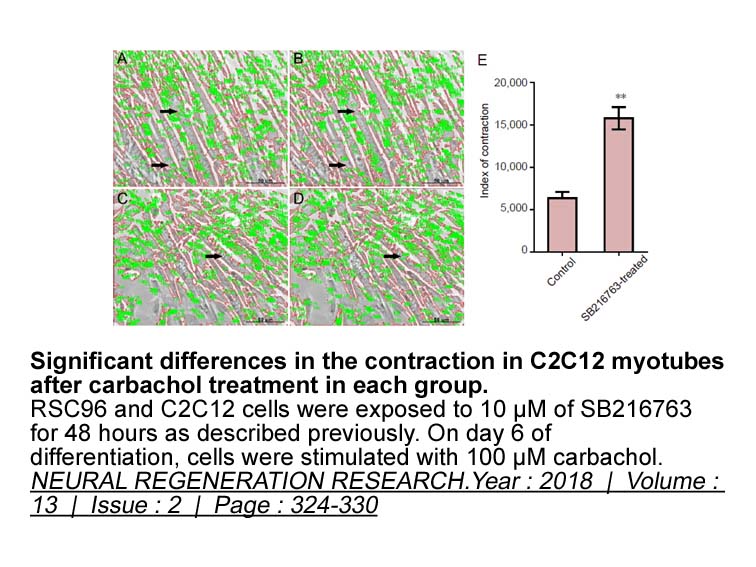
Transparency document Introduction Cigarette smoke is modifiable factors to alter the neurovascular functions. Now a day, cigarette smoking in public place is offense due to toxic exposure cause the neurological and respiratory system damage to non-smoker population [1]. A literature survey ex
-
The Kaempferia parviflora extract met the criteria for a
2019-10-21
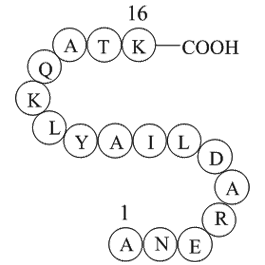
The Kaempferia parviflora extract met the criteria for a non-competitive interaction with CYP1A1 based on the decrease of Vmax and unchanged Km. The high Ki value and low IC50, as well as the high Km value was indicative of the weakness of the interaction between the Kaempferia parviflora extract an
-
br The ubiquitin proteasome system The ubiquitin proteasome
2019-10-21
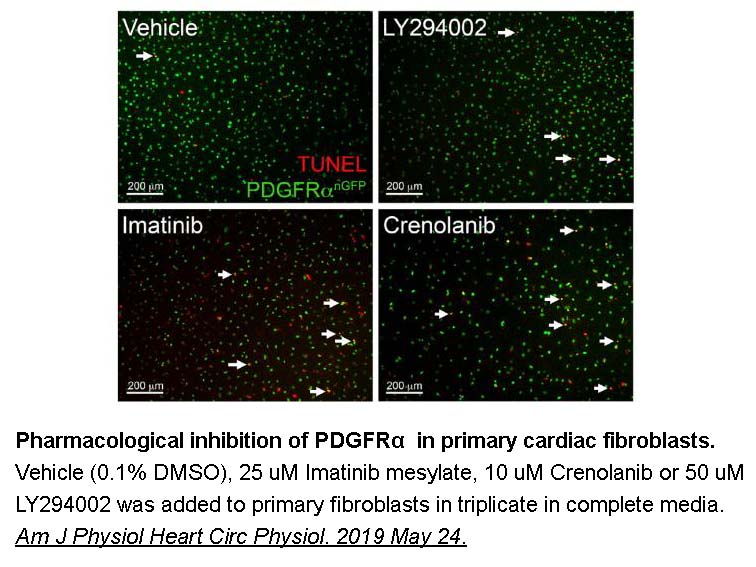
The ubiquitin proteasome system The ubiquitin proteasome system (UPS) plays a significant role in the regulation of cell growth and survival, in addition to maintaining cellular homeostasis. By means of the UPS, Roflumilast australia can precisely and temporally degrade approximately 80% of the
-
Immunofluorescence assays were performed to localize the enz
2019-10-21
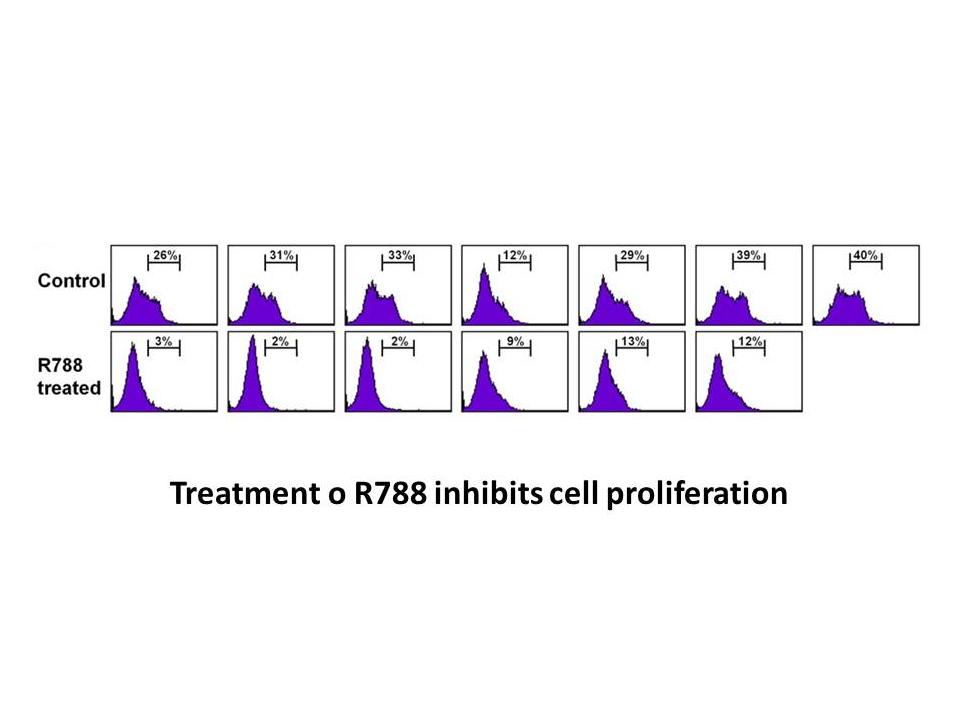
Immunofluorescence assays were performed to localize the enzyme during growth and differentiation to further characterize Giardia E1. As stated before, the assays showed that the Ebastine generated recognized specific E1 forms (E1-114, E1-90 and E1-67 with anti-gNTE1; and E1-47 with anti-gE1CT); th
-
br Materials and methods br Results br Discussion The
2019-10-21
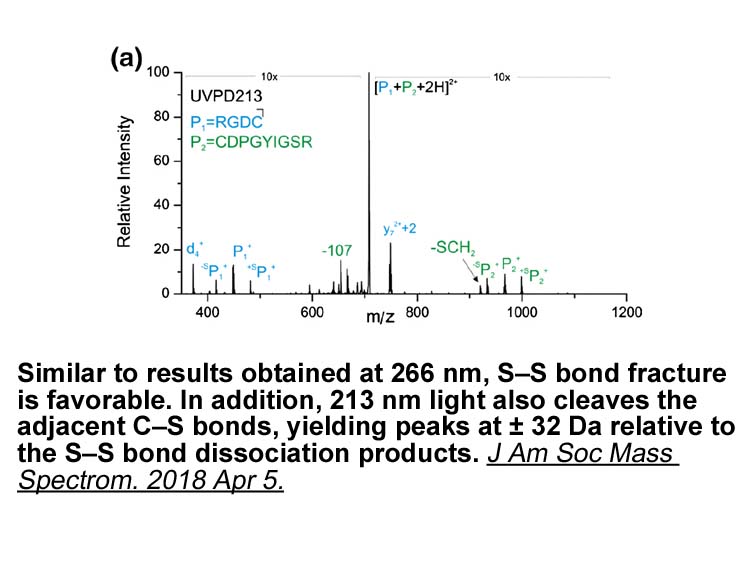
Materials and methods Results Discussion The incretin hormones potently stimulate insulin secretion, and it is generally assumed that this is mediated by an endocrine mechanism such that GIP and GLP-1 through the circulation reach the islet beta ic bupropion to stimulate their secretion. H
-
Tacrine amino tetrahydroacridine I Fig was the first AChEI
2019-10-19
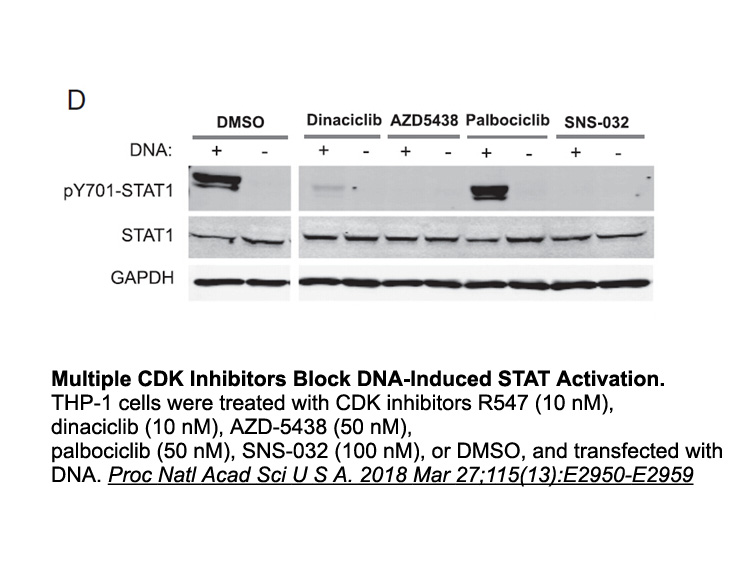
Tacrine (9-amino-1,2,3,4-tetrahydroacridine) (I, Fig. 1) was the first AChEI to be approved by the Food and Drug Administration (FDA) for the treatment of AD [25], [26], [27], [28]. However, despite its good AChE inhibitory activity, tacrine was far from ideal due to its low bioavailability and shor
-
Then the mechanisms responsible for BPA
2019-10-19
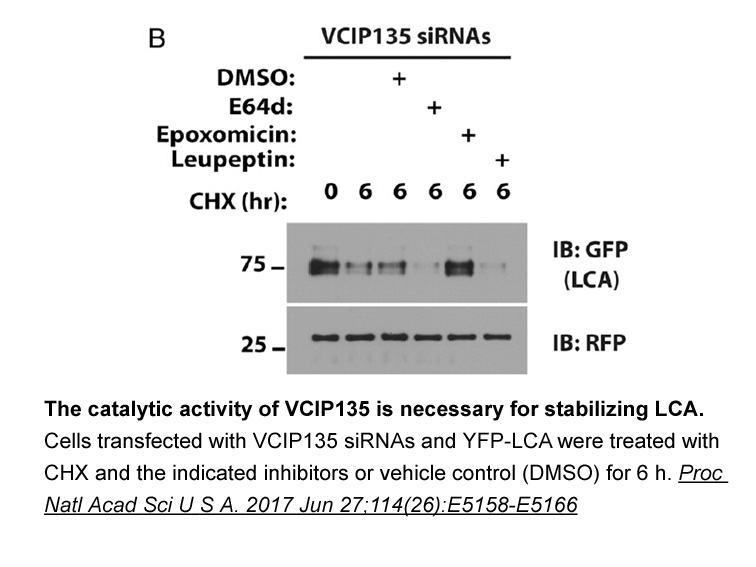
Then the mechanisms responsible for BPA induced up regulation of ERRγ were further investigated. Recent studies indicated that Akt and MPAK can modulate the protein levels and transcriptional activities of ERRγ (Heckler et al., 2014, Sun et al., 2014). Previous studies also indicated that BPA can ac
-
br Genes involved in oxysterol
2019-10-19
Genes involved in oxysterol metabolism The main enzymes that participate in the metabolism of oxysterols generally belong into one of two groups: transferases or oxidoreductases [22]. The following chapter describes those genes of oxysterol metabolism whose polymorphisms have been associated with
-
Because EBI is expressed on the
2019-10-19
Because EBI2 is expressed on the major subsets of immune cells, and small molecule antagonists for EBI2 were recently described (Benned-Jensen et al., 2013, Gessier et al., 2014), EBI2 constitutes a tempting drug target reminiscent of the sphingosine-1-phosphate receptor superagonist fingolimod/Gile
-
DGK is also involved in cell differentiation Previously to
2019-10-18
DGKδ is also involved in cell differentiation. Previously, to investigate the physiological roles of DGKδ, DGKδ-deficient mice have been generated. On the basis of the analysis of DGKδ-deficient mice, Crotty et al. reported that DGKδ regulates the epidermal growth factor receptor (EGFR) pathway by a
15932 records 962/1063 page Previous Next First page 上5页 961962963964965 下5页 Last page